INTRODUCTION
Chronic insomnia disorder (CID), a clinical condition, is characterized by the subjective experience of chronically disturbed sleep and sleep loss, and there is evidence of conditioned sleep difficulties and/or heightened arousal in bed [1]. These patients often present with difficulties falling asleep and maintaining sleep, and experience significant daytime consequences such as fatigue, sleepiness, and poor psychosocial function. In addition, patients with CID commonly have cognitive impairments and deficits in memory consolidation during sleep compared to that in good sleepers [2,3]. However, the neuroanatomical correlates of these symptoms and signs have not been clearly elucidated. Recent magnetic resonance imaging (MRI) studies providing a unique in vivo assessment of brain structural integrity have suggested that CID is associated with cortical and subcortical morphology alteration, which allow for an explanation of the characteristics of CID [4-9].
Brain morphometry using high resolution T1-weighted MRI has revealed anatomical brain changes associated with insomnia. Previous studies have considered the hippocampus as an important target of pathogenesis in CID [6,10,11]. A negative correlation between cognitive decline and hippocampal atrophy was reported in CID patients [9]. The frontal lobe and cingulate gyrus have been morphometrically evaluated in chronic insomnia [12]. Nevertheless, inhomogeneous patient brain compositions and different parameters and technologies had yielded inconsistent results across the studies. The current article summarizes the significant findings that localize cortical and subcortical changes associated with the characteristics of CID.
EXPLORING THE CORTEX
It is well-known that structural imaging may identify the anatomical substrates underlying disease-specific symptoms and signs in the field of neurology. Although MRI scans of individual patients with CID appear normal upon visual inspection, group analyses using diverse imaging techniques may reveal the specific changes in the brains. A number of morphometric studies were performed in insomnia patients using conventional high-resolution T1-weighted magnetic resonance images to quantify the size of specific brain structures. Among numerous methodologies, voxel-based morphometry (VBM) is the most popular automated technique as it provides a comprehensive assessment of anatomical differences throughout the brain [13,14]. The first optimized VBM study revealed that patients with CID had gray matter deficits in the left orbitofrontal cortex and precuneus compared to that of controls [15]. Optimized VBM has a significant circularity problem since the registration requires an initial tissue classification and vice versa [16]. In contrast, Statistical Parametric Mapping 8 (SPM8)-based VBM has updated a registration method termed Diffeomorphic Anatomical Registration Through Exponentiated Lie algebra, which is a more sensitive means of identifying differences in gray matter and white matter [16]. Thus, SPM8-based VBM provides more accurate localization than does optimized VBM in terms of supporting precise intersubject alignment and segmentation performance throughout the iterative unified model [16,17].
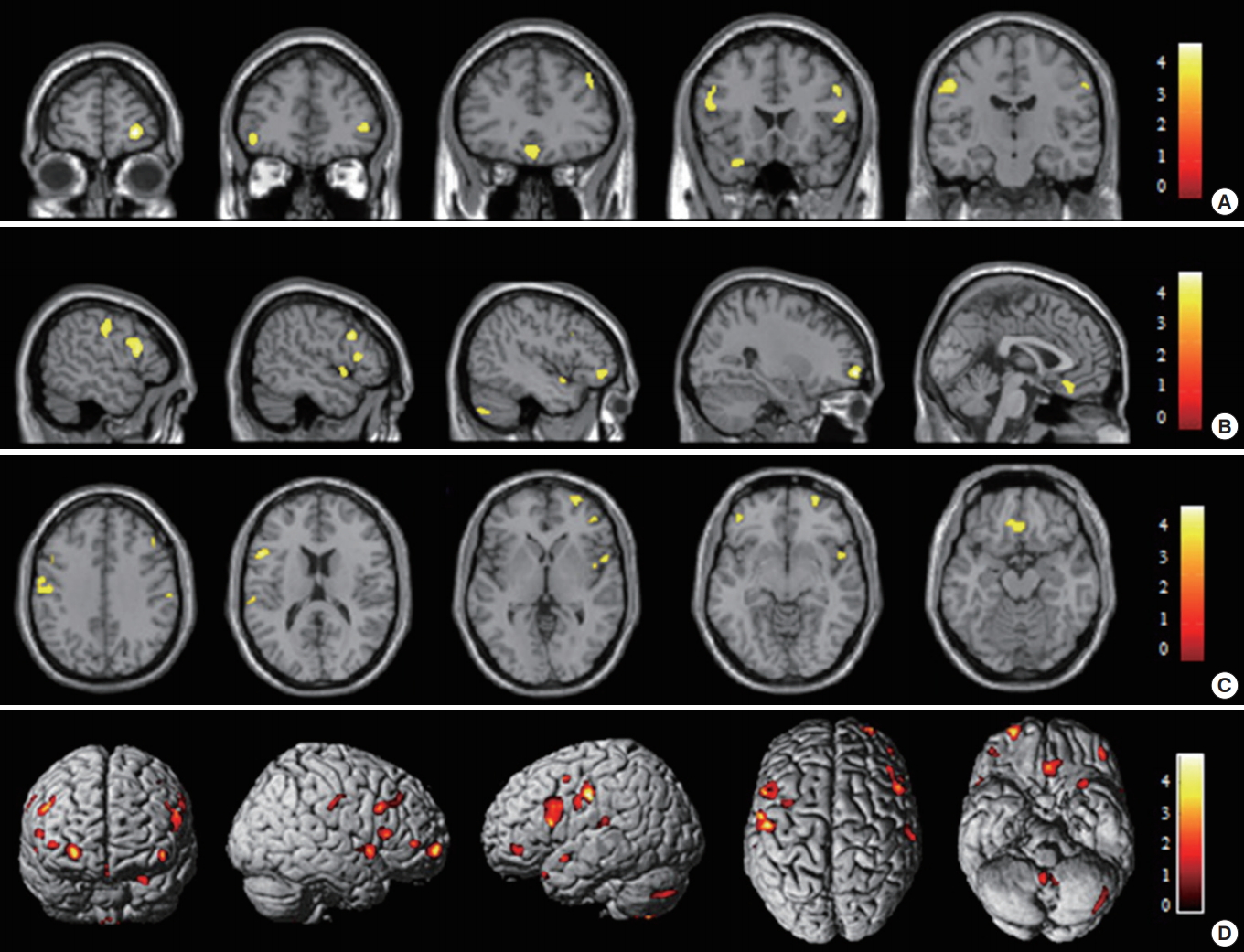
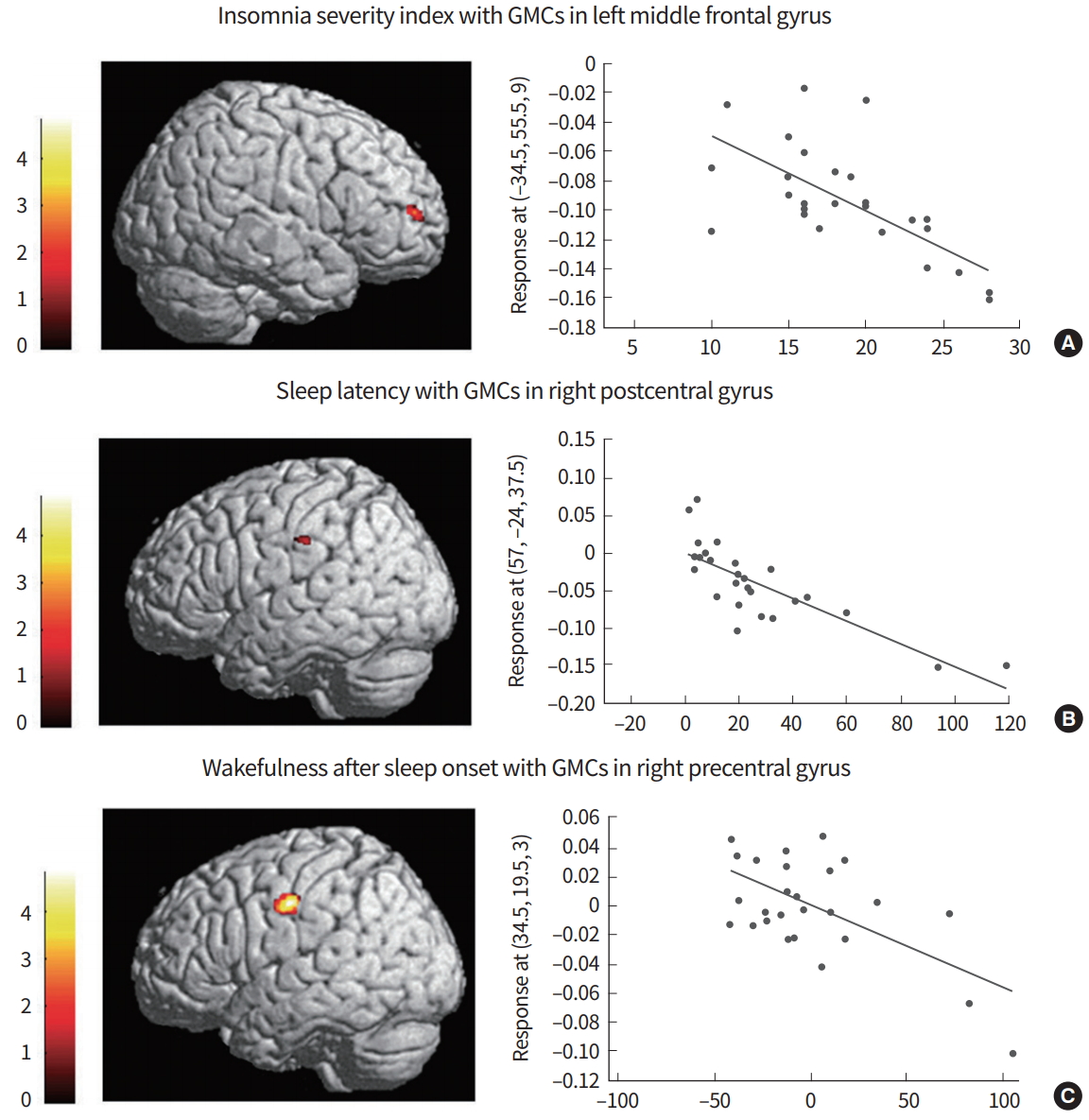
The first VBM study reported smaller gray matter volumes in the left orbitofrontal and parietal cortices in insomnia patients and a negative correlation between the orbitofrontal gray matter and insomnia severity, without any correlation with mood ratings [15]. A subsequent study [18] reported an increased volume of the rostral anterior cingulate cortex using FreeSurfer (Athinoula A. Martinos Center for Biomedical Imaging; http://www.freesurfer.net), an automated program for measuring the volume of brain structures [19]. However, this finding was not duplicated by one study using both the same methodology and VBM [6], and by two other studies using VBM [7,15]. Another study using the SPM8-based VBM method observed a significant reduction of the gray matter concentration in the left and right dorsolateral prefrontal cortices, pericentral cortex, and superior temporal gyrus compared to that in controls with a cluster threshold of 100 voxels at the level of uncorrected P<0.001. When a cluster threshold of more than 50 voxels was applied, the gray matter concentration was found to be decreased in the larger brain areas with the same anatomical coordinates as the results of the cluster threshold of 100 voxels, and was also observed to be decreased in the medial frontal gyri and cerebellum (Fig. 1) [5]. Gray matter deficits in the dorsolateral prefrontal cortex were related to attention deficit, frontal lobe dysfunction, and nonverbal memory decline in patients with CID, which might be associated with poorer sleep. Moreover, reduced gray matter concentrations in the left or right frontal cortices were significantly related to insomnia severity, longer sleep latency, or longer duration of wakefulness after sleep onset (WASO) (Fig. 2) [5]. This finding suggests that disturbed nocturnal sleep has a harmful effect on the frontal cortex of CID patients.
A previous VBM study identified the areas exhibiting only gray matter volume reduction in patients with chronic insomnia [15]. Thus, the two previous studies observed decreases in both gray matter concentrations and volumes, and the brain regions exhibiting volume changes were much smaller than those exhibiting concentration reduction. In optimized VBM, gray matter concentration in the local unit (i.e., voxel) can be transformed to the gray matter volume through the commonly known “modulation” process while accounting for regional stretching and compression occurring during coregistration [20]. The gray matter concentration is typically interpreted as gray matter tissue density relative to white matter, whereas gray matter volume is interpreted as absolute volume regardless of white matter [21]. Quantifying these two measures does not necessarily lead to overlapping results due to their different underlying properties but rather complements aspects of brain structural alterations [20].
Although it is unclear whether gray matter reductions are a preexisting abnormality or a consequence of insomnia, gray matter reduction as well as a lack of sleep or poorer sleep quality in the patients with insomnia might be responsible for the clinical features and cognitive dysfunction in chronic insomnia.
There has been an increase in the number of studies examining the corticocortical structural covariation network that is derived from correlational analysis of morphometrics between multiple cerebral regions [22-25]. Structural covariance may indicate altered neural connectivity resulting from relatively long-term processes such as neurodevelopment [26] or neurodegeneration [27]. Analyses of cortical thickness indeed demonstrated structural covariance within regions of the default mode network (DMN) with healthy aging [28]. In 60 selected patients with persistent insomnia symptoms from a population-based cohort study, significant cortical thinning was observed in both hemispheres of patients with insomnia compared to that in controls (left: 2.61±0.13 mm vs. 2.66±0.11 mm, P<0.05; right: 2.60±0.14 mm vs. 2.67±0.12 mm, P<0.01) [8]. Regional analyses revealed that cortical thinning was circumscribed to the left medial frontal cortex, bilateral precentral cortices, and right lateral prefrontal cortex, which was a similar pattern of gray matter decreases as that in the VBM study. Additional analysis of corticocortical morphological covariance demonstrated that insomnia patients only displayed a significant correlation of the medial frontal cortex with most of the frontal cortices, while high covariance (r>0.5, false discovery rate [FDR] <0.001) with the medial frontal cortex seed including the prefrontal cortex, precuneus, and lateral parietal cortex as well as scattered clusters in the temporal lobe was observed in controls. This analysis revealed that morphological alterations in the persistent insomnia group were circumscribed to multiple cortices, primarily the frontal and parietal cortices, and structural covariance was disrupted in the link between these two cortices in insomnia patients. This seems to be the most comprehensive structural imaging study of insomnia, demonstrating anatomical alterations and disrupted structural connectivity, as well as the implications on cognitive function and sleep quality. This study suggests that patients with persistent insomnia symptoms present with altered structural connectivity mainly within regions of the DMN that reduces their capacity to perform the normal transition to sleep, accompanied by a functional disconnection between the anterior and posterior regions of the DMN. This altered connectivity may further contribute to the sustained sleep difficulties and cognitive impairment commonly reported by insomnia patients [8].
EXPLORING THE SUBCORTICAL STRUCTURE
The negative association of disturbed sleep with performance on memory tasks [2,29,30] implicates hippocampal dysfunction in patients. Thus, there have been controversial findings concerning hippocampal volume in human insomnia studies. A previous pilot study found that the bilateral hippocampal volume was significantly lower in CID patients than in good sleepers [11]. In contrast, a recent study did not find any objective differences in the hippocampal volumes of CID patients, although some patients with sleep maintenance problems were found to have smaller hippocampal volumes, as determined by wrist actigraphy [18]. Although the authors found a significant correlation between reduced volumes in the bilateral hippocampi and poor sleep efficiency and increased WASO in those CID patients. The most recent study [4] reported that CID patients did not exhibit any definitive differences in intracranial volumes or in absolute and intracranial volumes to normalized hippocampal volumes compared to controls. However, a significant correlation was noted between the bilateral hippocampal volume and the duration of insomnia (left: r= –0.872, P<0.001; right: r= –0.868, P<0.001) and the arousal index of polysomnography (left: r= –0.435, P=0.045; right: r= –0.409, P=0.026) in patients [4]. In addition, they exhibited significantly impaired attention, frontal lobe function, and memory, and their verbal and nonverbal memory scores were positively correlated with the hippocampal volume. Another study did not reveal any statistical differences in hippocampal volumes between patients and controls [6]. These conflicting results regarding hippocampal volume may be related to the different anatomical landmarks to delineate the boundary of the hippocampus and different subsets of patients examined in the studies [4]. In particular, we observed that the left and right hippocampal volumes in insomnia patients were significantly and negatively correlated with the duration of insomnia and suggested that a longer duration of insomnia might negatively influence hippocampal function and volumes. These methodological or demographic inconsistencies always exist in neuroimaging studies, and well-standardized study protocols are required in multicenter trials to clarify the results.
Earlier hippocampal volumetry studies utilized manual delineation of the hippocampal boundary, which underlined the technical limitations of the low sensitivity of global hippocampal volumetry and the variability of hippocampal segmentation. To overcome these issues, automated subfield volumetry was developed and applied in the next study [7]. Vertex ( = point)-wise morphometry [31-33] based on a surface extracted from the manual segmentation of the whole hippocampus has been a surrogate to manual subfield volumetry. In 27 CID patients, a significant decrease was observed in the hippocampal volume compared to controls (left: 2,980±283 mm3 vs. 3,197±337 mm3; right: 3,079±298 mm3 vs. 3,247±404 mm3, P<0.05) [7]. This change was hemispherically symmetric; asymmetry between patients and controls did not differ. This finding was line with those of the manual hippocampal volumetry studies [15,18]. In patients, hippocampal atrophy was identified in all subfields. The largest cluster of atrophy was detected at the level of the hippocampal body and tail (FDR <0.005, 200 vertices) and was located medially, mainly within the region corresponding to the combined region of cornu ammonis (CA) 2-4 and the dentate gyrus (DG) (Fig. 3A) [7]. Atrophy at the level of the head was present on the superomedial surface corresponding to the CA1 region (FDR <0.05, 58 vertices). Hippocampal subfield atrophy in CID suggests reduced neurogenesis in the DG and neuronal loss in CA subfields in conditions of sleep fragmentation and the related chronic stress condition of insomnia. Atrophy in the CA3-4-DG region was associated with impaired cognitive function in patients (Fig. 3B), and these observations suggest that patients with chronic sleep disturbance are vulnerable to cognitive impairment.
A recent study exploring the morphological changes in subcortical structures demonstrated that local shape changes in the putamen were associated with higher arousal indices of polysomnography in CID [9]. In patients, atrophic changes in the hippocampus were associated with delayed correct response times in a Stroop word test, decreased phonemic word fluency in a Controlled Oral Word Association Test (COWAT), and lower Korean California Verbal Test (KCVLT) total scores. Amygdala atrophy was correlated with lower KCVLT short-delay free recall and recognition. Moreover, shape analysis of subcortical structures revealed that that lower sleep quality and a higher arousal index were associated with a greater number of atrophic changes in the hippocampus and putamen, and atrophic changes in the basal ganglia and thalamus were related to cognitive decline in neuropsychological domains (Fig. 4). These findings suggested that sleep disturbances in individuals with chronic insomnia are related to cognitive impairment consistent with the alteration of frontal-subcortical circuits. Additionally, this surface-based shape analysis method is useful in the localization of subcortical changes that are associated with cognitive decline in patients with CID.
CONCLUSION
Sophisticated neuroimaging techniques allow for the in vivo visualization of human brain anatomy with exquisite detail and the quantification of morphological changes. Comprehensive structural imaging studies demonstrated anatomical alterations in the frontal cortex, hippocampus, temporal cortex, or cingulate, and disrupted structural connectivity explaining the cognitive dysfunction and poor sleep quality of CID. Additionally, volumetry and subfield shape analysis identified atrophic changes in the hippocampus and putamen, which provided evidence for the pathophysiological mechanisms underlying the susceptibility of patients with CID to cognitive impairment. Advancements in imaging technology and software in larger, longitudinal studies may enable us to better understand CID and related disorders.